Orientation flights
Part of the reason for the success of honey bees is the division of labour between workers of different ages. Young workers (hive bees) clean the cells, nurse larvae and look after the queen. Older workers are foragers, collecting pollen and nectar (and water) from across the landscape.
To be successful, foragers need to know where to look and how to return.
The ‘where to look’ is partly accounted for by the well-known waggle dance {{1}}.
In this post I’m going to discuss the second component of successful foraging – the homing ability of foragers.
More specifically, I’m going to discuss how the bee first learns about the location of the hive.
Orientation flights
Bees do not instinctively know where the hive (or the tree they are nesting in for a wild colony) is located. They have to learn this before embarking on foraging trips to collect nectar or pollen.
This learning takes the form of one (or usually several – as we shall see) orientation flights. These enable the bee to memorise the precise location of the hive with relation to geographic landmarks. On subsequent foraging flights the bees use these landmarks to return to the hive.
Orientation flight have a characteristic appearance …
… and are very nicely described in the introduction to a paper by Capaldi and Dyer {{2}}:
An orientation flight at the nest entrance begins as a departing bee turns and hovers back and forth, turning in short arcs, apparently looking at the hive entrance. Then, the bee increases the size of the arcs until, after a few seconds, she flies in circles while ascending to heights of 5–10 metres above the ground. This spiraling flight takes the bee out of sight of human observers. She returns a few minutes later, always without nectar or pollen.
Which I couldn’t have written any better, so have reproduced verbatim.
There are a number of features of the orientation flight that are immediately obvious from this description (which all beekeepers will recognize). These include:
- A ‘local’ component, in the immediate vicinity of the hive
- Wider ranging flight at a greater altitude and a longer distance
- Direct observation does not allow the location, duration or track of these distant flights to be monitored
- The bee returning from the orientation flight does not bring pollen or nectar with her
Do orientation flights allow orientation?
How do we know that these flights enable the bee to learn where the hive is located?
Early studies conducted by Becker (1958) showed that bees captured after a single orientation flight and then re-released up to 700 metres away from the hive could find their way ‘home’. In contrast, bees that had not gone on an orientation flight before were, by definition, disoriented and did not return to the hive.
However, the percentage that returned after undertaking a single orientation flight was related to the distance of the release point, and was never more than ~60% (at 200 metres).
In contrast, reorienting older foragers (for example, as happens after moving a hive to a new location) were much better (~90%) at returning to the hive after a single reorientation flight.
Capaldi and Dyer extended these early studies by Becker to investigate the impact of the visibility of local landmarks on orientation and reorientation, and also measured the speed with which bees returned after being displaced.
Landmarks
These studies showed that a single orientation flight allowed bees to identify the landmarks in the immediate vicinity (100 – 200 m) of the hive. When released from more distant locations, returning flights were faster and more successful (i.e. fewer lost bees) when the bees had sight of the landmarks in the vicinity of the hive.
The hive itself was effectively invisible except at very short ranges. This makes sense for a tree-nesting animal. One tree looks much the same as another {{3}}, but if you learn that the nest is in the tree between the very tall conifer and the long straight hedgerow – two features visible from hundreds of metres distant – then orientation is straightforward.
This suggests that apiaries located near distinctive landscape features may be preferable in terms of increased returning forager rates.
“Distinctive” as far as an orienting worker bee is concerned, which may not be the same as distinctive to the beekeeper of course 😉
Reorienting bees (compared to first flight bees) took longer to explore the environment and were better at returning. Either these bees learn differently (a distinct possibility) or their prior experience in the wider landscape gives them an advantage when the hive is relocated.
Where do you go to my lovely?
The early studies by Becker and those by Dyer and colleagues defined many of the parameters that characterise orientation flights. What they did not do is show where the bees actually go during the orientation flights?
Do they just zoom around randomly?
Do they fly ever-increasing spirals?
Perhaps they perform some sort of grid search, exploring individual landscape features carefully for future reference?
Recent developments with harmonic radar have allowed tracking of individual bees during orientation flights over hundreds of metres. These have provided further insights into the process.
Because harmonic radar is also relevant to other studies of honey bee flight – for example, the impact of neonicotinoids on foraging ability – I’ll digress slightly from orientation flights to describe the technology.
Harmonic radar
Harmonic radar has revolutionised tracking studies of insects in much the same way as GPS tags have provided unique insights into bird migration (or, for that matter, shark migration).
The radar system has two components. The insect is tagged with a tiny antenna attached to a Schottky diode (together termed the transponder). The transmitter/detector is a ground-based scanner that transmits the radar signal. This is used as the energy source by the diode which re-emits a harmonic of the original signal which can then be detected.
Tagged bumble bee (left) and harmonic radar detector (right)
The transponder weighs less than a normal pollen load, though presumably there is some wind resistance from the antenna. In studies of bees with and without transponders fitted the orientation flights were of a similar duration, suggesting any wind resistance didn’t appreciably impact the flying ability of the bee.
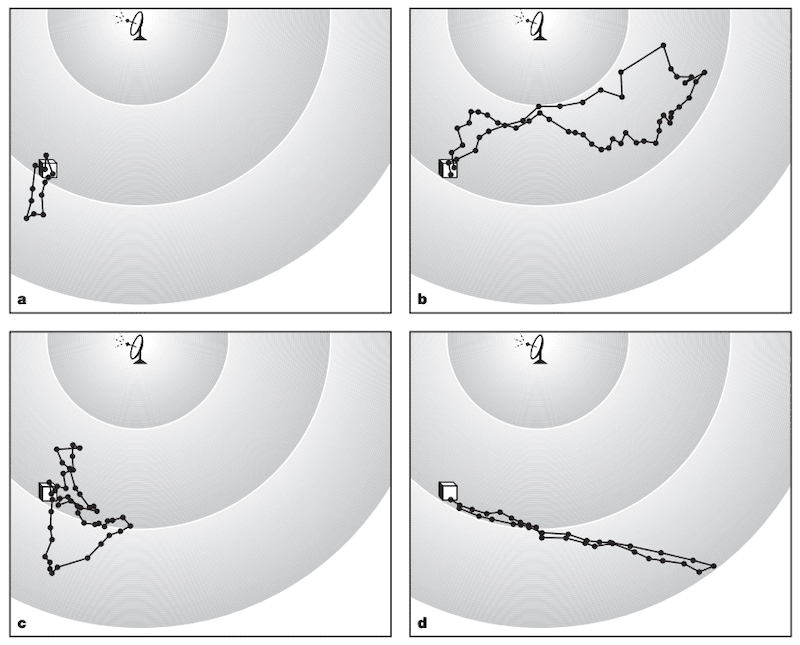
Orientation (a-c) and foraging (d) flights monitored by harmonic radar.
Orientation flights {{4}} were taken by bees between 3 and 14 days post-emergence, with the mean onset of foraging being 14 days post-emergence.
Bees took between 1 and 18 orientation flights, though there wasn’t a direct relationship between the number of flights and the age of the bee, suggesting they may learn at different rates.
Initial orientation flights were generally in the immediate vicinity of the hive. Older workers – pre-foraging – ventured further afield. More recent studies have addressed this in greater detail (see below).
Orientation flights were distinctly different from foraging flights. The former were slower and less direct. The ground speed of orienting bees was ~3.6 m/s in contrast to foragers who flew at ~5.6 m/s and, as shown in D above, foraging flights were very much ‘there and back’ straight lines.
Venturing forth …
A more recent study {{5}} has used harmonic radar to investigate multiple orientation flights by individual bees, effectively analysing how the bee explores the landscape as it ages towards a forager.
This was a remarkable study. It involved addition and subsequent removal of the transponder from 115 individual bees during 184 orientation flights. When the orienting bee returned they recaptured it, removed the transponder and allowed it to reenter the hive. When it reappeared for another orientation flight they reattached the transponder.
Anyone complaining about their inability to mark queens {{6}} should do this as a training exercise 😉
The scientists also recorded several foraging flights of a smaller number of the same bees, to allow comparison with their behaviour during orientation flights.
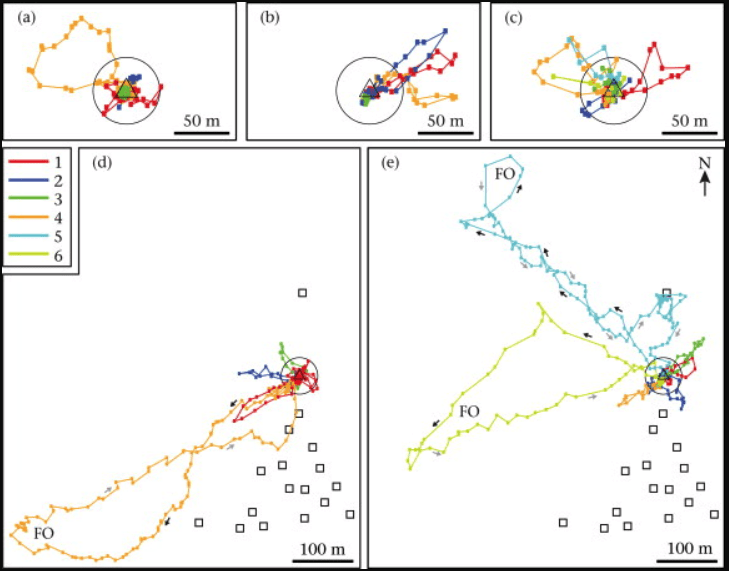
Orientation and foraging flights of five individual bees.
Flights were defined as short or long range, but long range orientation flights were still significantly shorter than foraging flights.
- Short range flights were made in poor weather and familiarised the bees with the immediate vicinity of the hive.
- Consecutive long range flights reduced in duration as the bees learnt about the immediate hive vicinity i.e. the long range flights included some local exploring at first as well.
- Orientation flights explored different areas of the landscape, rather than focusing on one sector.
- Subsequent foraging flights involved areas that the bees may have never visited during orientation flights.
- Some very long duration foraging flights may involve a degree of exploration, though it’s not clear whether this is truly orientation, or actual scouting activity.
Not all bees performed short and long range flights though early long range lights did involve local exploration as well.
Ground clues and conclusions
The final part of this study investigated the influence of visual landscape features on orientation flights. This deserves a post in its own right as the techniques are quite involved.
Essentially they generated heat maps of the flights overlaid onto the geography. Using this approach they determined that some features visible from the air e.g. borders between grassland and a track, influenced the direction of flight and hence the orientation flights.
There are additional studies of the influence of visible landmarks on bee flight which I’ll return to at some point in the future.
Again, like the comment made above about visible landmarks, it suggests to me that apiaries situated near such distinctive features may aid orientation and subsequent homing flights by honey bees.
When you next stand by your hive entrance on a warm, sunny afternoon and watch young workers flying to and fro across the entrance before spiralling up and away out of sight, you’ll know that it is an essential component of their training to be effective foragers.
They don’t forage for long – perhaps three weeks at most – but they are very effective, partly because they know where to return to.
Notes
Where do you go to my lovely? was the title of a rather syrupy (my blog, my opinion! … Apologies if it’s a favourite of yours 😉 ) song by Peter Sarstedt in 1969. It’s notable for some quite clever rhyming lyrics and a particularly dodgy mustache he sports in the YouTube video. (I’m just linking it, rather than embedding just because of the mustache).
It has nothing to do with bees.
{{1}}: I really should to write something about this …
{{2}}: Capaldi & Dyer (1999) The role of orientation flights in the homing ability of honey bees. J. Exp. Biol. 202:165-1666.
{{3}}: At least it does to me and, I’m assuming, to a bee.
{{4}}: See Capaldi et al., (2000) Ontogeny of orientation flight in the
honeybee revealed by harmonic radar. Nature 403:537-540.
{{5}}: Degen et al., 2015 https://doi.org/10.1016/j.anbehav.2014.12.030
{{6}}: Remembering that queens do not sting beekeepers.
Join the discussion ...