Dancing girls

In the 1967 film Doctor Doolittle Rex Harrison ended the Oscar-winning song 'Talk to the Animals' with the verse:
If we could talk to the animals, learn their languages
Think of all the things we could discuss
If we could walk with the animals, talk with the animals,
Grunt and squeak and squawk with the animals,
And they could squeak and squawk and speak and talk to us.
I talk to our dogs, but there's little evidence they understand anything I say (or perhaps they choose not to). Better trained dogs do understand dozens or even hundreds of commands, and can understand compound instructions, but that's still not a conversation.
In fairness, perhaps I'm at fault. I've usually no idea what our dogs are saying when they bark or growl, though I am able to distinguish between an angry bark and a happy bark.
Usually.
Almost all examples of human-animal communication involve very limited amounts of comprehension. We know that different populations of pygmy blue whales have distinct dialects and vocabularies, but we've no real idea what they are 'saying'. Similarly, we can discriminate between the alarm calls of monkeys advertising avian or terrestrial predators, and can elicit representative behavioural responses when we shout (in monkey-speak) 'snake' or 'eagle' at them.

For those readers not living where monkeys, eagles and snakes are endemic, the blue tit (Cyanistes caeruleus) also uses different alarm calls in response to a sparrowhawk or a cat.
A missed opportunity
Rex Harrison only mentions one insect in Talk to the Animals - the flea - and my Google searching suggests that studies of human-flea communication are few and far between {{1}}.
If we could talk to the animals, learn their languages
Maybe take an animal degree
I'd study Elephant and Eagle, Buffalo and Beagle
Alligator, Guinea Pig, and Flea
'Flea' was clearly included to rhyme with degree.
This seems like a missed opportunity.
'Bee' would rhyme just as well and - although we cannot truly communicate with them - we do understand some of the ways that they communicate with each other.
And we know what they're saying.
If we 'listen in' to what they're saying, we can learn useful things about the environment they are living in {{2}}.
The waggle dance
The communication I'm referring to is the waggle dance. This is a dance performed by worker bees in one of two circumstances and locations:
- By a forager on the face of the vertical comb within the nest. This dance is used to communicate the distance, bearing and quality of sources of forage - nectar or pollen - and water {{3}}. Other foragers are 'recruited' to better exploit the source, making foraging by these social insects much more efficient.
- By a scout bee on the surface of a bivouacked swarm. This dance is used to communicate the distance, bearing and quality of a potential nest site that the scout bee has located. The scout bee 'recruits' other scouts to check the nest site and these, in turn, dance to advertise it. Over repeated cycles of dancing and recruitment, 'better' sites are favoured and 'poorer' sites are forgotten, leading to a quorum decision being reached. The swarm then relocates, led there by the scout bees.
I've discussed the mechanics of the waggle dance before. Irrespective of whether the worker (scout or forager, these are dances only performed - or followed - by female workers, never by drones or queens) is dancing to indicate a great nest site, or a big patch of flowering fireweed, the essential components of the dance are the same.
Dancing in the dark
For convenience, I'll refer to the subject 'advertised' in the dance as the target.
Briefly, the dance consists of two phases:
- A figure of eight-shaped ‘return phase’ in which the bee circles back, alternately clockwise and anticlockwise, to the start of …
- The ‘waggle phase’, which is a short linear run in which the dancer vigorously waggles her abdomen from side to side.
The direction of the target is indicated by the angle of the waggle phase from gravity i.e. a vertical line down the face of the dance surface. This angle (α in the figure below) indicates the bearing from the direction of the sun that needs to be followed to reach the target.
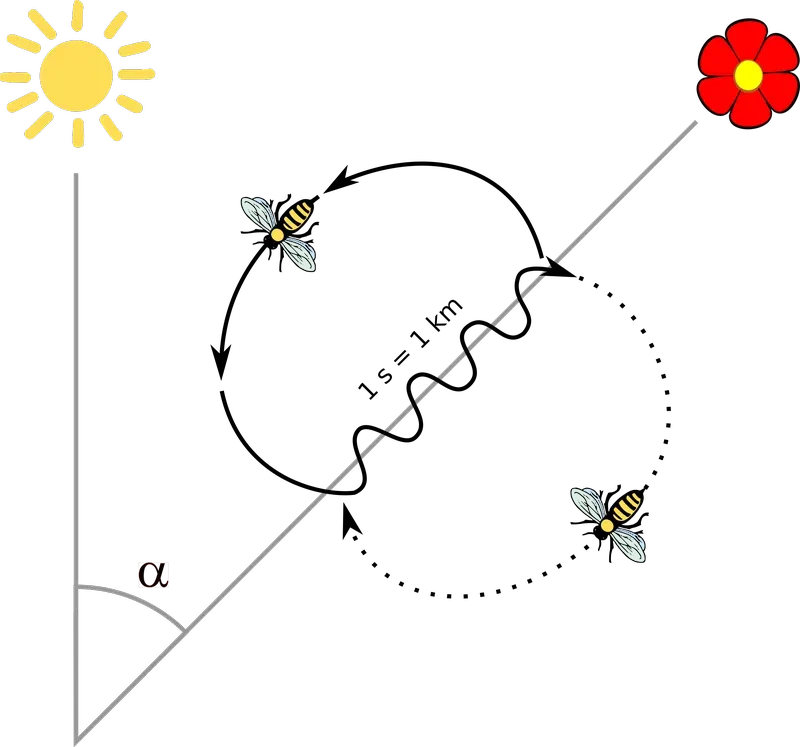
{kind=link}
Remember, at least within the nest, all of this occurs in the dark. The 'watching' bees that surround, and follow, the dance do so without receiving any visual cues {{4}}.
The waggle phase is the primary information content of the dance.
While the angle provides information on the bearing to be followed, the duration of the waggle phase provides information on the distance to the target. At a rough approximation, a one-second waggle phase indicates the target is one kilometre away (I'll return to this later as accurate calibration is influenced by the environment).
The quality of the target is communicated by the vigour with which the waggle is performed (and, additionally, by the speed with which the return phase is completed).
Limitations of the waggle dance
Karl von Frisch decoded the waggle dance and shared the 1973 Nobel Prize (with Nico Tinbergen and Konrad Lorenz) in Physiology and Medicine for his studies. It remains the only detailed informational communication by animals that we understand.
We cannot (yet?) directly ask bees questions that can be answered via the waggle dance.
However, that does not mean that we cannot use the waggle dance to answer questions we might have about honey bees.
In particular, if we know or control aspects of their environment - but want to know more - we can 'listen in' to the bees communicating with their nest mates and reach informed conclusions.
And that's what I'm going to discuss for the remainder of this post ... making interpretations about the environment, and the bees that occupy it, by reading the waggle dance.
The woods and the trees
Free-living honey bees build their nests in cavities that provide the space and protection the colony needs. In entirely natural environments these are largely - though not exclusively - in hollow trees.

All other things being equal - which, of course, they are not - this means that the density of honey bees should be higher in heavily forested areas, than treeless savannahs.
More trees = more cavities = more colonies.
One of those inequalities I hinted at is the availability of forage. If the forest has sufficient forage the statement above has more chance of being true. However, if the forage is insufficient - either throughout the season, or at particular times within the season - then colony numbers will be restricted to the carrying capacity of the environment.
Less forage = fewer colonies (irrespective of nest site availability).
Can we 'read' the waggle dance to make interpretations of the forage availability in forests?
We can, and I wouldn't have gone through that protracted introduction if it hadn't been done.
The results suggest that forage is limiting within forests - at least for honey bees - at certain times of the season. This has implications for free-living colony numbers in forests and, by extrapolation, for the management of forest environments to benefit honey bees and other pollinators.
Methodology
The study was relatively simple in concept. Hives were placed in different locations in managed forests and, periodically throughout the season, the waggle dances of bees were recorded as was the weight of the hive.
The weight indicated whether there was nectar available that could be stored. Interpretation of the waggle dance allowed the distance and direction to the source of the nectar or pollen being collected.
The important feature of the study was the choice of locations in which the hives were placed. Locations were chosen in areas with varying degrees of forest cover - from 50-99% - within 2 km (a typical maximum foraging distance).
Bees prefer to travel shorter distances to areas of forage of similar quality; this saves energy and reduces mortality when foraging. Therefore, if bees are observed to be travelling long-distances for forage, it means there is no equivalent or better source nearby.
The hives used were modified so that bees dancing on the outside of the outer frame could be observed and recorded. There's an assumption here - probably not unreasonable - that waggle dancing bees in that location of the hive are typical of bees dancing anywhere in the hive i.e. the position where the dance is conducted does not include additional informational content.
Hives were established in the forest location in July, treated for Varroa in August, and then data was recorded after overwintering between March and August i.e. the bees were fully acclimatised to their environment.
Dancing bees were assumed to be communicating pollen sources if their pollen baskets were filled, with the remainder indicating nectar or honeydew (or presumably water).
Six hours of recorded video was analysed for each day of recording, with ~25 dances/colony/day used for analysis after filtering out likely duplicate dances made by the same bee {{5}}. Over 2,000 waggle dances were analysed in total.
Calibration and wobbly waggles
The duration of the waggle phase is approximately 1 s/km. Bees don't measure distance per se, but instead measure optic flow - effectively the speed at which visual cues enter and leave their view - which is dependent upon the environment.
In complex environments there is a greater optic flow per unit distance. Conversely, in invariant environments, the optic flow is lower per unit distance.
Therefore, to accurately determine how far away the 'target' is that is being communicated in the waggle dance, the dance needs to be calibrated within that environment. This is achieved by siting a sucrose feeder at a known distance from the hive, recording the dances of bees reporting the location of the feeder and then accurately calculating the duration of the waggle phase per kilometre {{6}}.
The waggle dance is inherently error-prone, probably because the bees cannot dance any more accurately. This was taken into account during analysis when determining the preferred foraging locations.
Full details of the methods and interpretation can be found in the original paper (Rutschmann et al., 2023) which is freely available.
Forests are good ... some of the time
Of the ~2,000 dances analysed, ~75% were for nectar or honeydew with the remainder for pollen sources. Over 80% of the dances were for locations within 2 km of the study hive, with the mean foraging distance being ~1.25 km.
However, this season-long average hides a lot of month-to-month variation. The bees travelled much shorter distances in early Spring (~600 m) than midsummer (~1300 m) and usually needed to travel further for pollen than nectar/honeydew.
Forests appear to be more limited for pollen and less good in high summer for forage.
All of the above figures are averages over the 12 different regions. Foraging distances almost always increased in the more heavily forested regions.
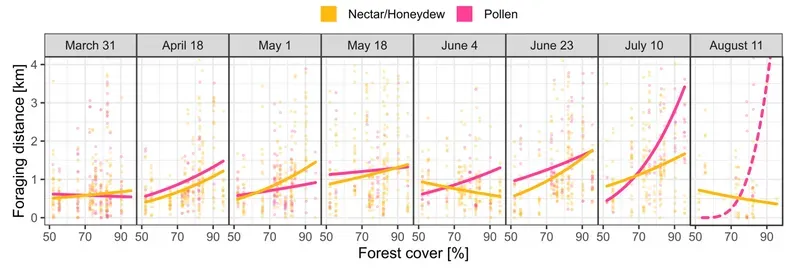
In the graphs above the lines indicate the average foraging distance for nectar or pollen in regions with different levels of forestation over the six months of analysis.
There were two exceptions to the 'more forest = greater distance' rule. In early June and mid-August, foraging distances for nectar/honeydew decreased with increasing forestation, indicating a rich source of these forage types during those periods.
By combining distance and bearing information from the dances the authors could determine whether the bees were foraging within the forest itself, or in cleared regions - clearings, rides, margins etc. - in the forest.
Foraging in cropland and grassland was always greater than predicted simply from habitat cover e.g. in >70% forest, the bees danced to indicate unforested regions 1.5-2 times as frequently as would be expected.
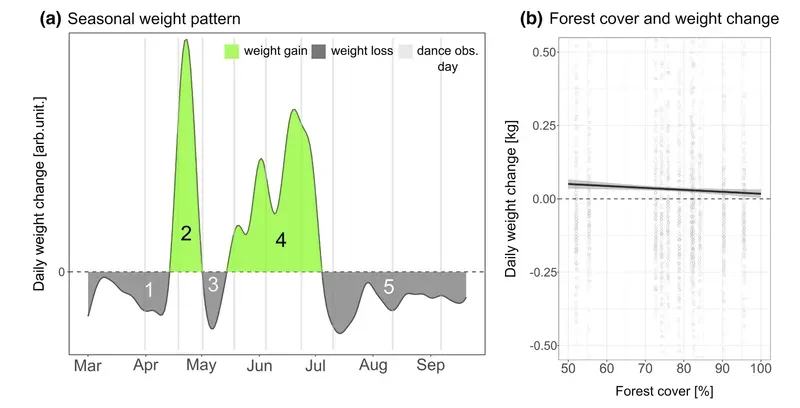
Finally, hive weight gain (normalised, not absolute) was unaffected by the level of forestation. Hives in forested regions still gained weight during Spring and mid-Summer, presumably as a consequence of the bees flying further for the nectar they were collecting {{7}}.
Managed forests and forest management
These studies were conducted in managed deciduous forests in Bavaria. The forests are predominantly beech (44%), with oak (21%), Scots pine (13%), spruce and hornbeam. As such they are very different from most of the managed forestry in the UK which is often dominated by invariant rows of conifers, usually Sitka spruce.

Honey bees forage within a very small region (geographically speaking) around the nest site/hive. This study shows that unbroken tracts of even deciduous forest are unsuited for managed - and therefore presumably free-living - honey bees.
That's perhaps not an unsurprising conclusion. Placing a colony in the centre of 125,000 acres {{8}} of oil seed rape would only benefit the colony when it was flowering. For the remainder of the season the colony would lose weight, or be forced to travel excessive distances to forage.
Honey bees do well in mixed environments where different forage types are available at different times throughout the season. It's worth bearing this in mind when choosing a location for a new apiary.
But what's true for honey bees is likely even more true for free-living solitary bees, and colony-forming bumblebees. The distances over which these bees forage is much more restricted (for example, analysis of 16 species of solitary bees demonstrated foraging ranges of 150-600 metres; Gathmann and Tscharntke, 2002), meaning that densely forested regions - even though they may be great at certain times during the season - are unlikely to be suitable habitats season-round.

Pollinator-friendly management of forests should include open spaces, rides and field margins.
Variety is the spice of life.
Nest site availability determined from the waggle dance
Free-living honey bees, and feral bees that have 'escaped' from hives due to inept swarm control, must find and occupy suitable nest sites to survive.
'Suitable' includes things like being sufficiently large to enable to colony to store enough honey to successfully overwinter, and protected - for example due to the height off the ground - so that they are not robbed out by a hungry bear {{9}}.
Eons of evolutionary time, coupled with selection by harsh winters or hungry bears, has honed the surveying capabilities of the scout bees that find new nest sites.
They only dance on the surface of the bivouacked swarm if the site they have found meets at least some of the criteria they (innately) look for, and they dance better i.e. more, and more vigorously, if the location meets more of the criteria {{10}}.
A small void with a large North-facing entrance at knee-height would elicit a much weaker (and so less convincing, and less likely to recruit followers) dance than one for a 40 litre void, 5 metres above ground with a small, South-facing, entrance.
I've discussed these requirements in posts on bait hives to attract swarms.
Lost and found
Tens of thousands of swarms are lost annually by beekeepers, but finding free-living colonies of bees - even in areas with lots of beekeepers - is relatively unusual.
Perhaps they all die out from disease during their first winter? Maybe they starve in their second season due to insufficient forage?
Alternatively, perhaps they never get established in a new nest site because these are limiting in the environment?

Some certainly die from disease, and some inevitably starve, but studies have shown that there are large numbers of free-living bees in the environment.
In Europe, estimates of the density of free-living and managed honey bee colonies are 0.3/km2 and 1.2/km2 respectively.
Clearly bees can survive, but could more survive if there were more nest sites available?
This question has recently been addressed by interpreting the waggle dances of scout bees on 'artificial' swarms.
The rationale is straightforward, if the number of potential nest sites reported exceeds the known density of free-living bees in the environment, then nest site availability is unlikely to be limiting.
Artificial swarms and 'artificial' swarms
To a beekeeper, an artificial swarm is a method of swarm control. For example, the Pagden method of swarm control, taught (often poorly {{11}}) in beekeeping courses all over the country, is often termed an artificial swarm.
However - as I described last year - it's not really very swarm-like at all. Please read that linked post for the gruesome details ... because I'm now going to confuse the matter further by describing a different type of artificial swarm.

One kilogram of worker bees (~5-10,000 bees) are removed from a colony, fed in the dark for 36 hours with as much thin syrup as they can consume, then allowed to cluster on a post to which a caged queen is attached. The post is fitted with a syrup feeder. The bees congregate around the caged queen and within an hour or so, scout bees can be observed dancing in the surface of the 'artificial' swarm.
The caging of the queen prevents the swarm relocating to a chosen nest site, and the dancing scouts are videoed for later decoding and analysis of the number, distance and bearing of selected nest sites.
These studies were conducted in East Sussex and are described in a recent paper by Visick et al., (2024). I've written about it as it's a good example of using our understanding of the waggle dance to answer questions about honey bees, even though we're unable to directly ask the bees a question.
Are nest sites limiting in natural environments?
In summary, no.
At least not in this study.
Using 14 'artificial' swarms in 2 different study areas (both containing mixed farmland, urban and woodland habitat), the authors analysed over 3,300 waggle dances. By setting up the 'artificial' swarms in different locations within the study areas they were able to survey acceptable (i.e. that induced scout bee dances) nest sites in overlapping regions spanning over 78 km2.
The chosen nest sites ranged from 0.1 km to over 11 km from the swarm location, with nest sites advertised in woodland (78% higher) and urban (43% higher) areas than farmland (12% lower) when compared with random predictions.
On average, swarms advertised over 5 nest sites (range 3-9) within the first ~15 hours of the swarm being created.
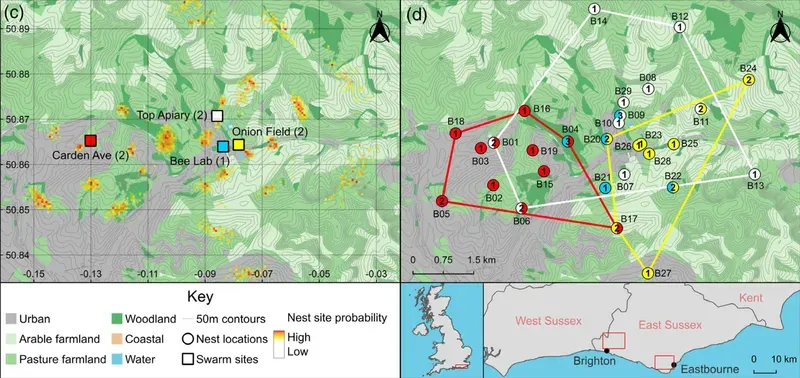
It's worth remembering this when considering what your bait hives are 'competing' with in the environment. The better you match the requirements of the scout bees, the more chance you will have of a swarm choosing your bait hive over the church tower, school entrance porch or ancient oak down the road.
Finally, by analysing the distance and bearing to the chosen nest sites, the density of potential nest sites in the environment could be predicted. In this study the estimated density of potential nest sites was 3/km2, a figure that exceeds the (unpublished) density of free-living bees in the study area (2/km2) and significantly higher than predicted density of 0.3/km2 in Europe.

In the environment in which this study was conducted - a mixed farmland/woodland/urban region of East Sussex - free-living honey bee numbers are not apparently limited by nest site availability.
How generally do these conclusions apply?
All environments are different, so it's not possible to conclude that natural nest sites are always available in excess.
The arable farmland my office window looks over now is dramatically different to the unmanaged (and unmanageable) mixed woodland it overlooked before I moved house.
However, it would be premature to conclude that the farmland has fewer nest site opportunities for the many any swarms I'm careless enough to lose.
The farmland is interspersed with small clumps of woodland, a few of which contain some venerable old trees (the largest circumference measured so far is about 6 metres). Old trees are more likely to contain suitable nest cavities for honey bees. In contrast, the unmanaged woodland on the remote west coast was relatively new growth, with few trees being even 1 metre in diameter.

Even ignoring forage availability and climate - both of which are wildly different in those two locations - I suspect natural nest sites are more frequent in the arable farmland interspersed with woods, abandoned coppices and old hedgerows ... to say nothing of the dilapidated farm outbuildings.
There are a number of interesting 'rewilding' projects with honey bees. These often involve adding new artificial nest sites into the environment. If these increase free-living bee numbers the conclusion would be that nest sites were previously limiting ... but it would be good to know this in advance.
If we had this information, either by direct measurement or extrapolation from studies such as those described above, it might be possible to focus rewilding efforts in areas where there is a paucity of natural sites, but that have ample year-round forage.
However, as shown by the first study ... suitable nest sites are, alone, insufficient. Forage (and climate, and a host of other factors) must also be suitable. With good environmental conditions the impact of disease - and disease resistance - becomes important, and more readily determined.
To be continued ...
Sponsor The Apiarist
The Apiarist covers 'the science, art and practice of sustainable beekeeping ... so much more than honey' and only exists because of the countless hours I spend talking to myself in the car and then failing to remember the beautifully crafted sentences when faced with a keyboard and screen.
If this, or other, posts has helped or inspired your beekeeping, or was amusing or enlightening, then please consider sponsoring The Apiarist.
Sponsorship costs less than £1/week annually, or little more than a large cappucino monthly. Sponsors help ensure the weekly posts appear, receive an irregular (approximately monthly) newsletter, and have access to an increasing number of sponsor-only content ... those starred ⭐ in the lists of posts.
Alternatively, you can help reduce my caffeine overdraft ... and please spread the word to encourage other beekeepers (or non-beekeepers ... the more the merrier) to subscribe.
Thank you.
References
Gathmann, A., and Tscharntke, T. (2002) Foraging ranges of solitary bees. Journal of Animal Ecology 71: 757–764 https://onlinelibrary.wiley.com/doi/abs/10.1046/j.1365-2656.2002.00641.x.
O’Regan, H.J. (2018) The presence of the brown bear Ursus arctos in Holocene Britain: a review of the evidence. Mammal Review 48: 229–244 https://onlinelibrary.wiley.com/doi/abs/10.1111/mam.12127.
Rutschmann, B., Kohl, P.L., and Steffan-Dewenter, I. (2023) Foraging distances, habitat preferences and seasonal colony performance of honeybees in Central European forest landscapes. Journal of Applied Ecology 60: 1056–1066 https://onlinelibrary.wiley.com/doi/abs/10.1111/1365-2664.14389.
Visick, O.D., Adams, I., Ney, P., Marzano, F.S., and Ratnieks, F.L.W. Do nest sites limit wild honey bee colonies? Decoding swarm waggle dances to assess nest site availability. Ecological Entomology n/a https://onlinelibrary.wiley.com/doi/abs/10.1111/een.13361.
{{1}}: I'll let someone else write the grant application, but I'd appreciate some credit for highlighting this research opportunity.
{{2}}: And probably other things as well ... yet to be determined.
{{3}}: I don't think they communicate the quality of water, but the sentence was starting to get convoluted, so I lumped them all together.
{{4}}: Is there visual information conveyed by scouts dancing on the surface of the bivouacked swarm? I don't know.
{{5}}: So avoiding pseudo-replication within the dataset.
{{6}}: Both studies discussed in this post used relevant calibrated waggle dances.
{{7}}: But did they gain enough weight to overwinter?
{{8}}: The approximate area within 2 km.
{{9}}: Although the chances of this happening in East Sussex are limited, it wasn't all that long ago - in evolutionary terms - that bears roamed South-East England (O'Regan, 2018).
{{10}}: I'm interested in whether there is a precedence in what they look for ... is void volume more important than height above the ground?
{{11}}: Been there, done that.
Join the discussion ...